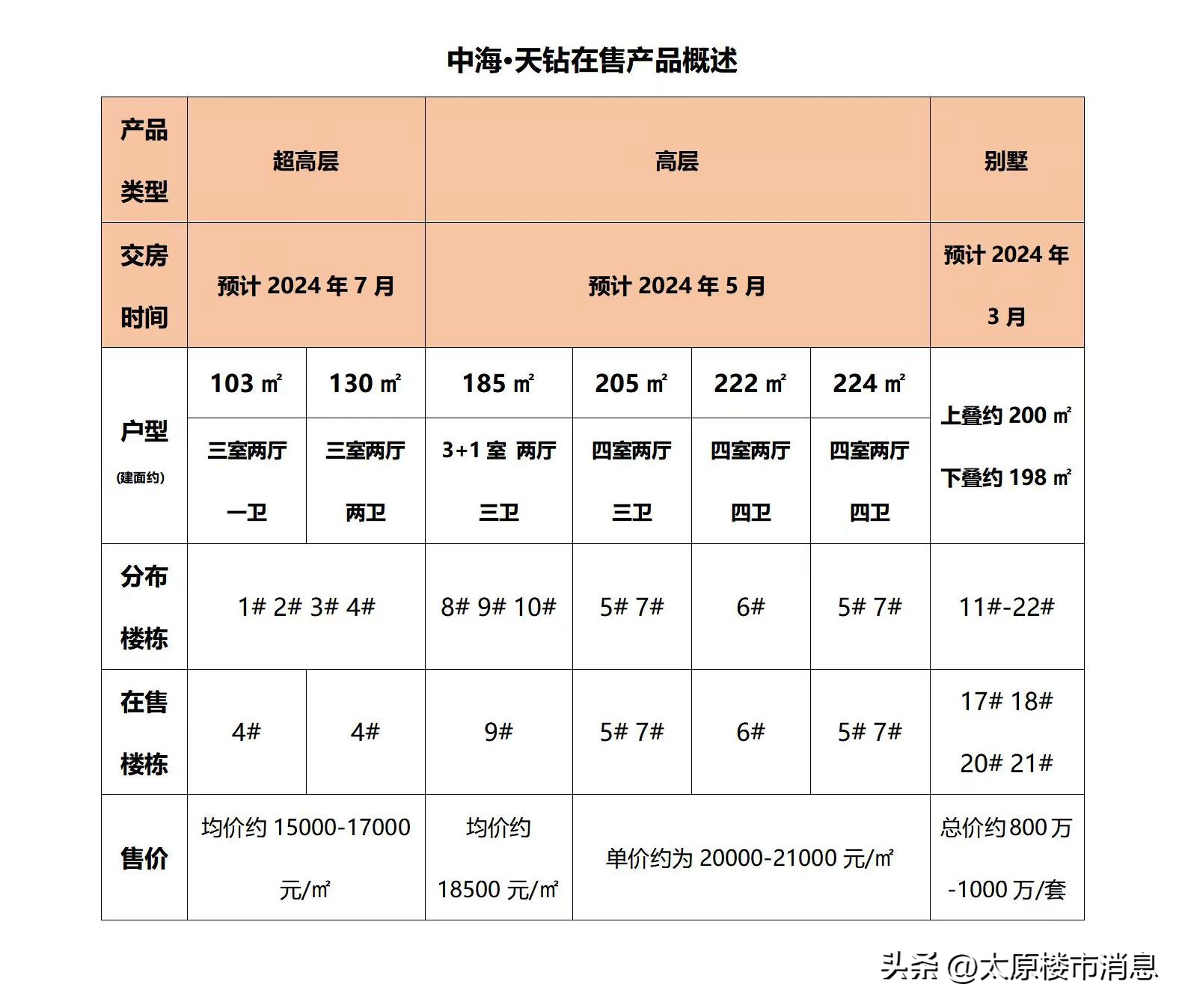
预计气候变化将导致全球平均气温上升,高纬度地区气温上升幅度更大,极端温度事件的频率也会增加。温度变化会导致生理、形态、生活史和物种分布的变化。考虑到这一点,温度变化对物种的影响是并且将是广泛的,对群落和生态系统具有潜在的级联效应。因此,迫切需要了解物种如何对温度变化做出反应。

为了了解和预测气候变化对群落的影响,重要的是要确定影响物种对温度敏感性的特征。这些包括形态特征,例如大小或颜色,以及生理和行为特征,并且可能是物种栖息的热环境的结果。在全球范围内,温带和热带地区之间的热环境之间存在重大差异,热带地区的温度范围比温带地区更窄,年平均温度更高预测,与温带地区相比,热带地区较低的温度变化可能会选择热带物种之间更窄的热耐受性(“季节性”假说)。

因此,尽管预计高纬度地区的变暖程度更大,但热带物种可能比温带物种更容易受到温度变化的影响,因为它们应对变化的能力较差。此外,热带物种比温带物种更接近其热最大值,因此可能更容易受到相同水平的变暖的影响。因此,温度的微小变化将对热带物种的生存和健康产生严重后果。然而,其他研究表明,一些温带物种可能同样容易受到气候变化的影响。

例如,活动时间短的温带物种可能只会经历狭窄的温度范围,并且可能以与热带物种类似的方式选择低热耐受性。 尽管在热带地区发现了超过50%的昆虫物种,但缺乏系统的实地研究,调查热带变温动物的体温调节能力,并比较热带和温带物种。目前尚不清楚来自不同生态背景和纬度的物种对气候变化的敏感性差异程度。

气候变化对物种的影响通常在宏观气候尺度上进行测试,使用粗尺度的气象站数据,假设物种生活在这些环境温度下。这没有考虑到个别物种在异质环境中经历的小规模微气候温度差异,也没有考虑到在这些环境中移动一小段距离的小型变温动物会导致暴露于非常不同的气候条件。事实上,小气候和气象站之间的温度可能有很大差异,气象站将热带林下森林的年平均温度高估了0.5°C,高地高估了高达2.0°C。

小规模的温度变化可以在物种的热适应中发挥重要作用,并且可以缓冲或放大温度升高的影响。我们缺乏关于宏观气候尺度上温度上升如何影响小气候温度的知识,以及物种如何受到这些细微的温度尺度差异的影响。蝴蝶作为指示性分类群、传粉媒介、猎物和食草动物很重要,但作为变温动物可能特别容易受到温度变化的影响。了解它们如何应对温度变化对于评估气候变化对整个生态系统的影响至关重要。

蝴蝶可以通过多种方式调节体温,包括在不同的生命阶段或一天中的不同时间选择有利的热微气候。他们还使用其他行为机制,例如翅膀相对于太阳的方向,来增加或减少热吸收。形态特征,包括翅膀颜色、反射率、大小和头发长度,在确定体温调节能力方面也很重要。例如,浅色蝴蝶可以通过用翅膀将太阳辐射反射到胸部以加热或返回环境中以冷却来调节体温。深色蝴蝶比浅色蝴蝶升温更快,这也许解释了为什么它们在凉爽的气候中更频繁地被发现,例如森林内部或高纬度地区,能够快速升温可以提高健康。

大小也可以决定体温调节能力,同源山蝶的温度调节在栖息地利用和体型方面因物种而异。大型物种可以利用翅膀的表面积来拦截阳光,并且通常比小型物种更擅长体温调节。最后,蝴蝶家族之间的体温调节能力存在系统性差异,与其他温带家族相比,温带Pieridae物种特别擅长体温调节。在这项研究中,我们比较了来自新热带(巴拿马)和温带地区的蝴蝶种群的缓冲能力-独立于周围气温调节体温的能力。

我们假设(H1)来自温带地区的蝴蝶,自然暴露在更大的温度范围下,将比来自新热带地区的蝴蝶能够更好地缓冲体温对空气温度变化的影响。我们在组合和家庭层面对此进行了测试。由于在我们的欧洲站点和巴拿马站点发现的蝴蝶物种存在显着重叠,我们将它们视为单个组合,从而产生了每个地区最常见和最引人注目的蝴蝶的温带类群和新热带类群。
如果观察到新热带和温带地区之间缓冲能力的差异,我们假设(H2)这将由蝴蝶在不同地区暴露的温度驱动,而不是由其形态特征的差异驱动。最后,我们假设(H3)温带蝴蝶作为对温带气候的适应,会主动选择比新热带蝴蝶更温暖的小气候,并且它们会比新热带物种更多地通过姿势手段升温。

方法
遇到蝴蝶时(不追逐)用蝴蝶网捕获蝴蝶,使用的协议收集数据,如下所示。一旦进入网中,并在10秒内,蝴蝶胸部的温度读数使用热电偶(直径0.5毫米)和手持式指示器拍摄。空气温度是在捕捉蝴蝶的腰部高度拍摄的,热电偶遮挡了太阳。如果蝴蝶在捕获前停留在基板上,则用热电偶记录其坐姿上方1厘米的空气温度(微气候温度,Tm)。蝴蝶被鉴定为物种或亚种。对于来自热带Calephelis属的蝴蝶,不可能识别个体到物种,因此这些蝴蝶的数据被汇总到属级别。
使用卡尺测量从翅膀尖端到与胸部相交点的前翅长度(以毫米为单位)。我们根据巴拿马和英国每个物种的现场数据计算了平均前翅长度。除了有现场数据的Erebia spp外,捷克共和国和奥地利蝴蝶的平均前翅长度取自文献。翅膀长宽比(强烈影响飞行性能,这反过来又可以决定物种移动到更有利的小气候的能力,从而决定它们缓冲空气温度的能力。

所有新热带和温带物种的翅膀长宽比是根据文献中的照片计算的。为此,每个物种的五个雌性和五个雄性镶嵌标本的照片被获取并上传到ImageJ。测量前翅长度(如上)和深度(前翅前缘和后缘之间的最长线,垂直于翅长轴测量),并用于计算每个标本的前翅长宽比。然后计算每个物种的平均纵横比。对于有性二态物种,雌性和雄性的平均前翅长宽比分别计算。
统计分析
为了确保我们对缓冲能力的估计是稳健的,只有至少10吨的物种b在至少5°C的 T 范围内进行测量自被纳入分析。为了估计每个物种的缓冲能力,使用T进行简单的线性回归b作为响应变量和 T自因为预测变量是为每个物种单独拟合的。提取关系的斜率,从中减去以提供缓冲能力的估计值(黄油能力估计值)。使用这种方法,陡峭的斜率表明蝴蝶的 Tb在小范围的T 上变化很大自因此表明缓冲能力低。浅坡度显示蝴蝶的 Tb在较宽的 T 范围内保持相对稳定自从而表明具有很高的缓冲能力。

在我们的研究区域共采集了六个科(Hesperiidae,Lycaenidae,Nymphalidae,Papilionidae,Pieridae和Riodinidae)的94种(新热带:54种,n=1333;温带:40种,n =5482)。在欧洲只发现了一种Riodinidae物种(Hamearislucina),因此我们对这个家族的分析专门比较了该物种与其新热带亲戚的缓冲能力。罂粟科物种仅在新热带地区取样,因此在比较新热带和温带地区科内的缓冲能力时,该科被移除。
为了测试新热带和温带地区蝴蝶组合之间以及每个蝴蝶家族内缓冲能力的差异,分别拟合了组合和每个家族的线性混合效应模型。拟合体温作为响应变量,T自、区域(温带或新热带)及其双向相互作用作为预测变量。物种作为随机效应被包括在内。进行似然比检验以确定具有双向交互作用的模型是否比没有项的模型拟合得更好。如果是这样,则表明两个区域中的蝴蝶组合或家族的缓冲能力不同。如上所述,提取并减去每个区域(新热带和温带)的斜率估计值,以获得每个区域每个组合和家庭的平均缓冲能力估计值。

结果
从新热带地区,测量了来自1333个物种的54个个体。空气温度(T自)温度范围为17.4°C至39.7°C,平均为28.5°C。体温(Tb)温度范围为22.1°C至40.3°C,平均为31.7°C。从温带地区测量了5482个物种的40个个体(结果S1)。T自温度范围为10.0°C至34.8°C,平均为22.2°C。b温度范围为15.4°C至45.0°C,平均为28.0°C。新热带物种的缓冲能力估计范围为-0.317(Hemiargushanno)至1.005(Phoebisargante),平均值为0.295。温带物种的缓冲能力估计范围为-0.404(Hamearislucina)至0.675(Erebiamedusa),平均值为0.243。
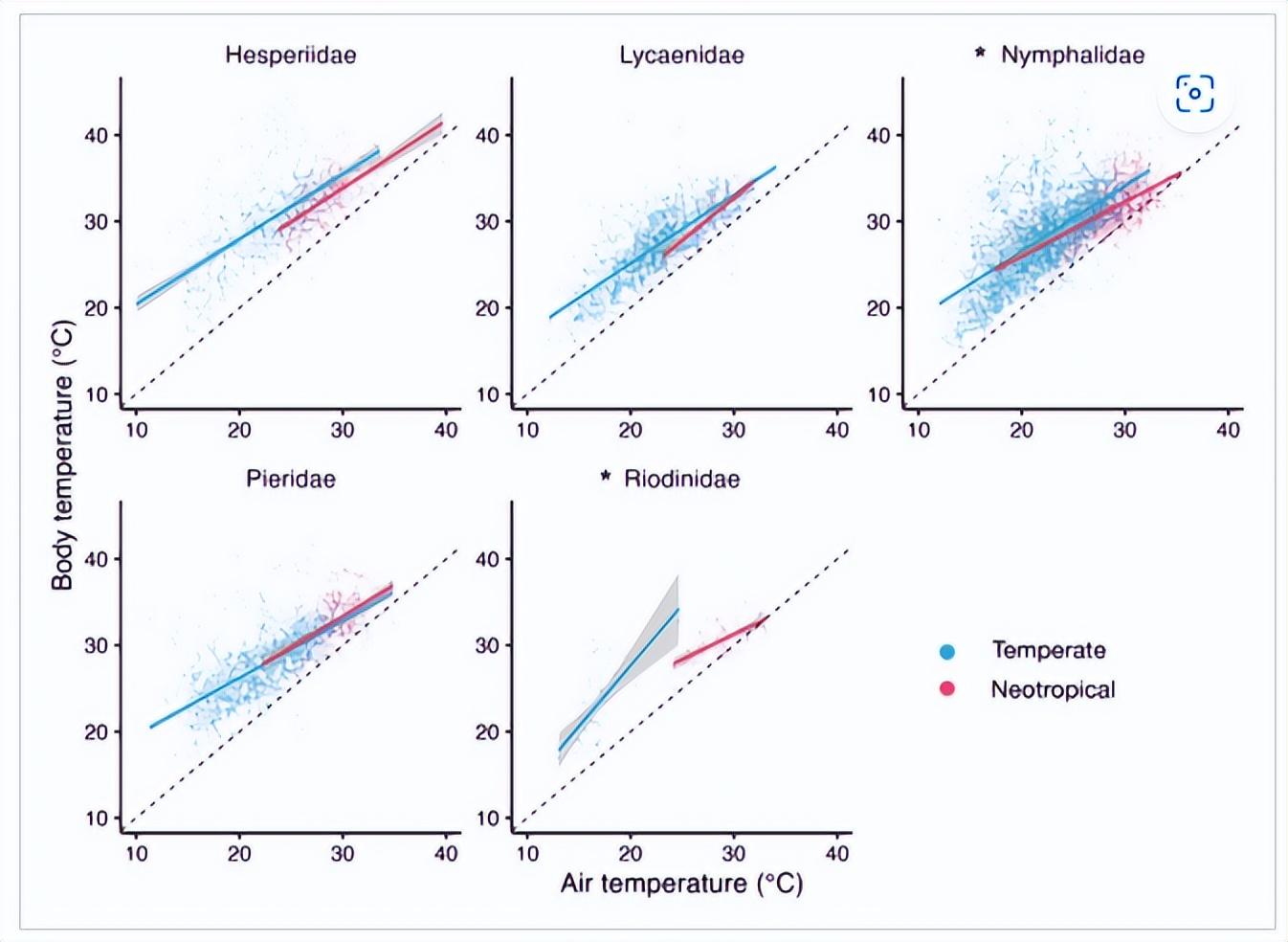
温带和新热带蝴蝶组合的缓冲能力是否不同?
在整个组合中,新热带蝴蝶的平均缓冲能力(0.350±0.035)显著高于温带蝴蝶的平均缓冲能力(0.220±0.010),新热带物种似乎能够更好地降低其Tb在更高的 T自 (χ2=14.17,df=1, p <.001;图1)。这意味着在20.0°C的 T 范围内自,预测新热带蝴蝶Tb将变化13.0°C,而预测的温带蝴蝶Tb将变化15.6°C。当数据限制在T范围内时,也获得了类似的结果。自这发生在两个地区。

结论
据我们所知,这是第一项比较热带和温带系统之间变温空间的热缓冲能力的研究。通过使用标准化方法来测量缓冲能力和机制,我们提供了一个模板,用于在分类群中进行跨纬度比较,从而增加对跨纬度物种如何应对温度变化的理解。我们强调需要进一步的跨纬度研究,比较热带和温带物种的热能力。
尽管世界上一半以上的昆虫物种生活在热带系统中,但大多数研究都集中在温带变温动物的热能力上。未来的研究应侧重于热带变温动物,并扩大对这些物种如何调节温度及其对气候变化的潜在脆弱性的知识。我们还强调了在预测热带和温带物种对未来气候变化的脆弱性时考虑物种形态特征以及使用小气候选择和姿势温度调节的重要性。
未来的研究还应调查栖息地结构在蝴蝶缓冲能力中的作用,例如确定有利的小气候如何成为热带和温带物种极端温度的避难所,为保护物种免受变暖的管理方案铺平道路。最重要的是,我们的结果表明,单个物种具有自己独特的体温调节能力。我们强调,在仅根据物种所居住的热环境对气候变化的脆弱性做出普遍假设时,需要谨慎。

引用
1.Advani,N.K.,Parmesan,C.和Singer,M.C.(2019)。来自纬度和海拔范围限制的Melitaeacinxia蝴蝶的起飞温度:对太阳辐照度的潜在适应。生态昆虫学,44,389-396。
2.Bates,D., Mächler,M.,Bolker,B.,& Walker,S. (2015). 使用lme4拟合线性混合效应模型。统计软件杂志,67,1-48。
3.Bonebrake,T.C., Ponisio,L.C., Boggs,C.L.,& Ehrlich,P.R. (2010).不仅仅是指标:热带蝴蝶生态和保护综述。生物保护,143,1831-1841。